
1. Erreger der Lyme-Borreliose
Der Erreger der Lyme-Borreliose wurde 1982 von Willy Burgdorfer entdeckt und 1984 als neue Borrelienart Borrelia burgdorferi klassifiziert. Bakterien des Genus Borrelia gehören zur Ordnung Spirochaetales (Spirochäten), denen eine einzigartige Zellanatomie gemeinsam ist: Sie haben sogenannte Endoflagellen, die unter der äußeren Membran liegen und subterminal an den beiden Enden des Protoplasma-Zylinders inserieren.

Abbildung 1: Zellanatomie und Querschnitt der Borrelia Burgdorferi. Aus Rössle, Wanner, Wilke, 2001.
 Dies befähigt sie, schraubenförmige Bewegungen auszuführen und sich so effektiv in hochviskösen Medien (z. B. extrazelluläre Matrix) fortzubewegen. B. burgdorferi ist ca. 20 µm lang und nur ca. 0,3 µm dick (Borrelia burgdorferi) und lichtmikroskopisch im Dunkelfeld oder mit Spezialfärbungen wie der Fuchsinfärbung darstellbar.
Dies befähigt sie, schraubenförmige Bewegungen auszuführen und sich so effektiv in hochviskösen Medien (z. B. extrazelluläre Matrix) fortzubewegen. B. burgdorferi ist ca. 20 µm lang und nur ca. 0,3 µm dick (Borrelia burgdorferi) und lichtmikroskopisch im Dunkelfeld oder mit Spezialfärbungen wie der Fuchsinfärbung darstellbar.
 Der Erreger der Lyme-Borreliose wurde zunächst als einheitliche Art angesehen. Erste Hinweise auf eine größere Heterogenität des Erregers der Lyme-Borreliose lieferte die immunologische Analyse europäischer Isolate. Aufgrund molekulargenetischer Analysen wurden bisher 10 Genospezies beschrieben, wovon bisher drei Arten als sicher humanpathogen gelten: B. burgdorferi sensu stricto (s.s.), B. garinii und B. afzelii. B. valaisiana wird als weitere humanpathogene Spezies diskutiert.
Der Erreger der Lyme-Borreliose wurde zunächst als einheitliche Art angesehen. Erste Hinweise auf eine größere Heterogenität des Erregers der Lyme-Borreliose lieferte die immunologische Analyse europäischer Isolate. Aufgrund molekulargenetischer Analysen wurden bisher 10 Genospezies beschrieben, wovon bisher drei Arten als sicher humanpathogen gelten: B. burgdorferi sensu stricto (s.s.), B. garinii und B. afzelii. B. valaisiana wird als weitere humanpathogene Spezies diskutiert.
Geografische Verteilung der B. burgdorferi sensu lato Spezies
- B. Burgdorferi sensu stricto: Nordamerika, Europa
- B. garinii: Eurasien
- B. afzelii: Eurasien
- B. lusitanae Europa, Nordafrika
- B. valaisiana: Eurasien
- B. bissettii: USA
- B. andersonii: USA
- B. japonica: Japan
- B. tanukii: Japan
- B. turdi: Japan
Fettdruck: humanpathogen
Als Oberbegriff der verschiedenen Genospezies wurde B. burgdorferi sensu lato (s.l.) vorgeschlagen. Bisher konnten B. afzelii und B. garinii nicht in den USA und B. burgdorferi s.s. nicht in Japan isoliert werden. B. valaisiana wurde bisher nur in Europa nachgewiesen. Aufgrund der Heterogenität des äußeren Membranproteins OspA wurden mindestens 7 verschiedene OspA-Serotypen definiert, die eng mit der genetischen Spezies-Klassifikation korrelieren. OspA-Serotyp 1 enspricht der Spezies B. burgdorferi s.s., OspA-Serotyp 2 der Spezies B. afzelii und die OspA-Serotypen 3-7 der Spezies B. garinii.
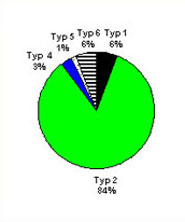
Verteilung von OspA-Serotypen bei europäischen B. burgdorferi sensu lato Stämmen
Liquor (n=43)

Zecken (n=90)

Haut (n=68)
| Liquor | Zecken | Haut | |
|---|---|---|---|
| B. Burgdorferi | 19 % | 20 % | 6 % |
| B. afzelii | 12 % | 9 % | 84 % |
| B. garinii | 69 % | 71 % | 10 % |
Nach Wilske, 2001.
Die Verteilung der einzelnen OspA-Serotypen und somit auch der Spezies ist bei Haut-, Liquor- und Zecken-Isolaten unterschiedlich. Die Spezies B. afzelii wird ganz überwiegend bei Hautisolaten gefunden, während Isolate von Zecken und humane Liquorisolate heterogen sind. B. garinii wurde am häufigsten aus Liquor isoliert. Mittels OspA-Typ-spezifischer PCR wurde in Gelenkpunktaten von Patienten mit Lyme-Arthritis nachgewiesen, dass auch die Erreger der Lyme-Arthritis heterogen sind. Dies impliziert, dass verschiedene OspA-Serotypen unterschiedlichen Organotropismus und/oder unterschiedliche Disseminationsfähigkeit in Zecke oder Patient aufweisen. Ein weiteres wichtiges Oberflächenprotein OspC (siehe Kapitel Impfung und Diagnostik) weist noch eine deutlich größere Heterogenität als OspA auf.
Verteilung der OspA-Typen bei der Lyme-Arthritis
20 PCR positive Patienten von 31 Patienten mit Lyme Arthritis; ein Patient hatte eine Doppelinfektion mit Typ 4 und 5
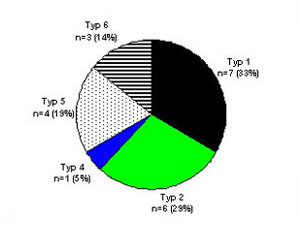
- B. burgdorferi s.s.: n=7 (33 %)
- B. afzelii: n=6 (29 %)
- B. garinii: n=8 (38 %)
- Nach Wilske, Herzer, Effert, 2001.